... Phases Magazine N° 22
Des nanodisques ultrarigides de
tensioactifs
MARS 2000 N° 23
Une équipe du Spht (Physique Théorique) applique des méthodes de physique statistique à un modèle de protéine qui rend compte du caractère universel des structures secondaires (hélices et feuillets).
Le repliement des protéines est un des problèmes majeurs de la biologie moléculaire, schématiquement décrit comme suit (voir encart). Chaque protéine est un petit polymère qui est capable d'exister sous deux formes, l'une dépliée (ou dénaturée) et biologiquement inactive, l'autre repliée (ou native) et biologiquement active. Comprendre le repliement, c'est comprendre le lien entre la suite des acides aminés le long de la séquence primaire de la protéine et la forme repliée responsable de l'activité biologique.
|
Depuis une dizaine d'années, les physiciens théoriciens s'intéressent à ce problème. Si on oublie le solvant, la physique du repliement apparaît comme une physique d'amas avec contrainte polymérique. Une description plus chimique d'une protéine souligne alors le contraste entre la grande variété des acides aminés, et l'aspect (presque) périodique de la chaîne principale ( - CONH - Ca - ).
Ces remarques suggèrent deux modèles extrêmes possibles pour une protéine :
un modèle qui tient compte de la structure spécifique de chaque protéine. On décrira alors la protéine comme un polymère où chaque monomère est différent (hétéropolymère).
un modèle où l'on s'attache à décrire certains aspects universels des protéines comme l'existence de régions spatialement structurées de la chaîne principale (en hélice ou en feuillet). La protéine est alors décrite comme un polymère où tous les maillons sont identiques (homopolymère).
|
|
Une équipe du SPhT a récemment étudié un modèle qui va au-delà du modèle simple d'homopolymère : tous les acides aminés sont identiques à l'exception de la glycine et de la proline. Ses caractéristiques majeures sont (i) la présence d'un fort moment dipolaire électrique dans le plan peptidique CONH, (ii) l'existence d'une même chiralité pour les Ca, (iii) des contraintes stéréochimiques fortes (impliquant par exemple que le moment dipolaire du plan CONH est perpendiculaire au vecteur joignant les deux Ca adjacents).
Dans cette approche, la glycine apparaît comme un défaut de chiralité, et la proline comme un défaut de moment dipolaire. En effet, on remarque expérimentalement que glycines et prolines sont très souvent situées hors des hélices et des feuillets.
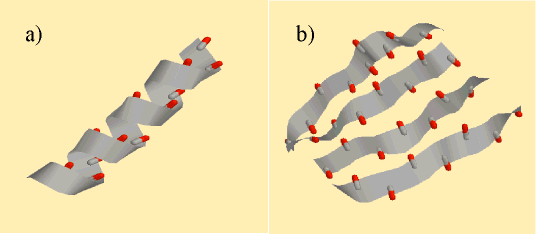
Ce modèle, traité au niveau du champ moyen, présente une analogie formelle avec des transitions de phase des cristaux liquides. On trouve ainsi, pour une chaîne dipolaire assez chirale, une transition entre une phase dépliée et une phase "cristalline". Le paramètre d'ordre associé à cette transition est un tenseur qui adopte la même valeur pour les hélices et les feuillets (Fig. 1). Cette unité des structures secondaires permet de comprendre l'existence de séquences "caméléons", qui, pour une même structure primaire, adoptent une conformation soit en hélices, soit en feuillets, suivant leur place dans la protéine.
Le modèle d'une chaîne dipolaire chirale a permis d'expliquer l'existence des structures secondaires, et peut être appliqué à l'étude des mécanismes du repliement. Cependant la détermination de la structure repliée d'une protéine particulière, ainsi que la description de sa dynamique de repliement ou de son activité biologique, devra prendre en compte les aspects hétéropolymériques.
Pour en savoir plus :
Protein folding, anisotropic collapse and blue phases,
E. Pitard, T. Garel and H. Orland,
J. Phys. I France 7 (1997) 1201.
Contact :
E. Pitard, T. Garel, H. Orland.
Phases Magazine N° 23
Des nanocristaux de silicium dans l'espace !...
 Les Protéines
Les Protéines